ДНК – единственные молекулы, способные к самовоспроизведению. В синтетический период интерфазы митотического цикла перед каждым делением клетки происходит удвоение молекул ДНК (синтез ДНК или репликация ДНК), благодаря этому и осуществляется передача генетической информации от материнской клетки к дочерним.
ДНК-репликация (от лат. репликацио – свернуть, отвернуть) – самоудвоение молекулы ДНК. При ДНК-репликации двойная спираль молекулы ДНК сначала разделяется на 2 полинуклеотидные цепи, затем на каждой из образовавшихся цепей из свободных нуклеотидов интерфазного ядра в соответствии с правилом комплементарности азотистых оснований достраиваются дополняющие дочерние цепи. Каждая вновь образовавшаяся молекула ДНК, таким образом, состоит из одной материнской полинуклеотидной нити и комплементарной ей дочерней нити. Процесс ДНК-репликации лежит в основе автодубликации хромосом.

Рис. 31. Репликация ДНК
Репликация ДНК (рис. 31) – процесс матричный. Так как две цепочки молекулы ДНК комплементарны друг другу (т.е. азотистые основания дополняют друг друга), то каждая из них может служить шаблоном или матрицей для построения новых цепей.
Для удвоения ДНК необходимы:
- материал, из которого будут строиться новые молекулы ДНК;
- энергия, т.е. молекулы АТФ;
- ферменты;
- матрица – основа, по которой строятся новые полинуклеотидные цепочки.
Главными "действующими лицами" репликации являются ферменты ДНК-полимеразы. Связано это с особенностью фермента ДНК-полимеразы, участвующей в реакции полимеризации дезоксирибонуклеотидов, и с тем, что две цепи молекулы ДНК антипараллельны. Впервые это было обнаружено в 1957 г., когда А. Корнберг обнаружил у кишечной палочки фермент, катализирующий процесс полимеризации ДНК из нуклеотидов; он был назван ДНК-полимеразой. Затем ДНК-полимеразы выявили и в других организмах. Было показано, что субстратами всех этих ферментов служат дезоксирибонуклеозидтрифосфаты (дНТФ), полимеризующиеся на одноцепочечной ДНК-матрице. ДНК-полимеразы последовательно наращивают одноцепочечную цепь ДНК, шаг за шагом присоединяя к ней следующие звенья в направлении от 5'-концу к 3'-концу, причем выбор очередного дНТФ диктуется матрицей.
Репликация ДНК – процесс сложный, требующий точности и быстроты, что достигается благодаря участию в процессе целого комплекса ферментов. Кроме ДНК-полимеразы в процессе синтеза ДНК принимают участие и другие ферменты, например ДНК-лигазы, которые сшивают дезоксирибонуклеотиды в цепочку; группы ферментов, раскручивающих спираль ДНК перед репликацией и группа ферментов, закручивающих вновь синтезируемые молекулы дочерних ДНК в спираль.
Таким образом, в результате репликации по программе исходной материнской ДНК образуются две дочерние молекулы, идентичные материнской. Каждая вновь сформировавшаяся молекула состоит из одной полинуклеотидной цепи материнской молекулы и одной полинуклеотидной цепи – вновь синтезированной. Такую репликацию называют полуконсервативной. Такой механизм репликации был экспериментально доказан в 1958 г. М. Мезельсоном и Ф. Сталь.
Поскольку синтез ДНК происходит на одноцепочечной матрице, ему должно предшествовать обязательное разделение (хотя бы на время) двух цепей ДНК. Исследования, проведенные в начале 60-х гг. на реплицирующихся хромосомах, выявили особую, четко ограниченную область репликации, перемещающуюся вдоль родительской спирали ДНК и характеризующуюся местным расхождением двух ее цепей. Эта активная область из-за своей Y-образной формы была названа репликационной вилкой. Именно в ней ДНК-полимеразы синтезируют дочерние молекулы ДНК.

Рис. 32. Схема строения репликационной вилки ДНК
(О.О. Фаворова, 1996)

Рис. 34а. Репликация ДНК прокариот
(http://www.college.ru/biology/)

Рис. 34б. Репликация ДНК вирусов
(http://www.college.ru/biology/)

Рис. 34в. Репликация ДНК эукариот
(http://www.college.ru/biology/)
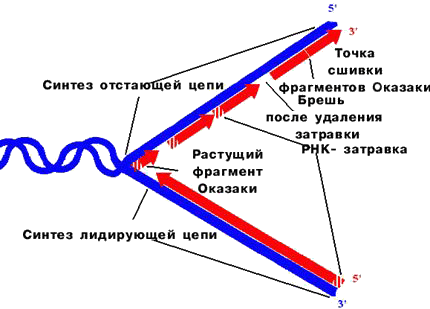
Рис. 33. Прерывистый синтез ДНК
Легко вообразить, что репликация происходит путем непрерывного роста нуклеотида за нуклеотидом обеих новых цепей по мере перемещения репликационной вилки; при этом, поскольку две цепи в спирали ДНК антипараллельны, одна из дочерних цепей должна была бы расти в направлении 5'–3', а другая в направлении 3'–5'. В действительности, однако, оказалось, что дочерние цепи растут только в направлении 5'–3', то есть всегда удлиняется 3'-конец затравки, а матрица считывается ДНК-полимеразой в направлении 3'–5'. Это утверждение, на первый взгляд, кажется несовместимым с движением репликационной вилки в одном направлении, сопровождающимся одновременным считыванием двух антипараллельных нитей. Разгадка секрета заключается в том, что синтез ДНК происходит непрерывно только на одной из матричных цепей. На второй матричной цепи ДНК синтезируется сравнительно короткими фрагментами (длиной от 100 до 1000 нуклеотидов, в зависимости от вида), названными по имени обнаружившего их ученого фрагментами Оказаки. Вновь образованная цепь, которая синтезируется непрерывно, называется ведущей, а другая, собираемая из фрагментов Оказаки, – отстающей. Синтез каждого из этих фрагментов начинается с РНК-затравки. Через некоторое время РНК-затравки удаляются, бреши застраиваются ДНК-полимеразой и фрагменты сшиваются в одну непрерывную цепь ДНК специальным ферментом. Это так называемый прерывистый синтез ДНК.
Неспособность реплицировать ДНК – летальное событие для любой клетки.
ДНК бактерий, митохондрий, хлоропластов и некоторых вирусов представляет собой кольцевую двойную спираль. Молекулы ДНК сохраняют во время репликации кольцевую форму, и репликация такого кольца проходит одновременно в двух направлениях, т.е. существуют две репликативные вилки. Обе точки возникают в одной точке и удаляются от нее в обоих направлениях, пока снова не встретятся в точке терминации. Репликация состоит из большого числа последовательных этапов, которые включают узнавание точки репликации, расплетание родительской двухцепочечной молекулы, удержание ее цепей на достаточном расстоянии друг от друга, инициацию синтеза новых дочерних цепей, их удлинение (элонгацию), закручивание цепей в спираль и терминацию репликации. Все эти этапы процесса репликации протекают с большой скоростью и исключительной точностью.
Репликация эукариотической ДНК, которая упакована в нуклеосомы и находится в составе хроматиновых волокон, осуществляется гораздо сложнее, чем репликация бактериальной ДНК. Эукариотическая ДНК тоже реплицируется в двух противоположных направлениях, но репликативные вилки двигаются на порядок быстрее. Потребовалось бы слишком много времени для репликации эукариотического генома, который намного больше бактериального, если в каждой хромосоме была бы только одна точка репликации, поэтому их много (порядка тысячи). Репликация хромосомной ДНК начинается сразу во многих точках начала репликации, и из каждой такой точки одновременно в противоположных направлениях движутся две репликативные вилки. Точки начала репликации расположены группами по 20–80, называемыми репликативными единицами. Все эукариотические хромосомы реплицируются одновременно, и в ядре эукариотической клетки работает одновременно много тысяч репликативных вилок. В ходе репликации активируются новые репликативные единицы, пока не будет реплицирована вся ДНК. Каждая точка начала репликации должна быть использована только один раз во время репликации хромосомы, поэтому с использованными точками начала связываются белки, предотвращающие повторную инициацию репликации в этом участке.