Французские исследователи Франсуа Жакоб и Жак Моно в 1961 г. опубликовали разработанную ими модель негативной индукции на примере регуляции экспрессии лактозного оперона. За своё открытие спустя 4 года они были удостоены звания нобелевских лауреатов.
Лактозный оперон находится под постоянным негативным воздействием, вызванным ассоциацией его области оператора со специфическим белком-репрессором (рис. 12, А). Особенность этого белка заключается в том, что он имеет две специфические области узнавания молекул лактозы. Если лактоза присутствует в клетке, то она взаимодействует с белком-репрессором (по две молекулы лактозы на одну белковую молекулу репрессора), что приводит к изменению его пространственной укладки (конформации) и отсоединению от оператора (рис. 12, Б). Происходит индукция лактозного оперона. Таким образом, лактоза сама выступает в качестве индуктора синтеза ферментов своего катаболизма.
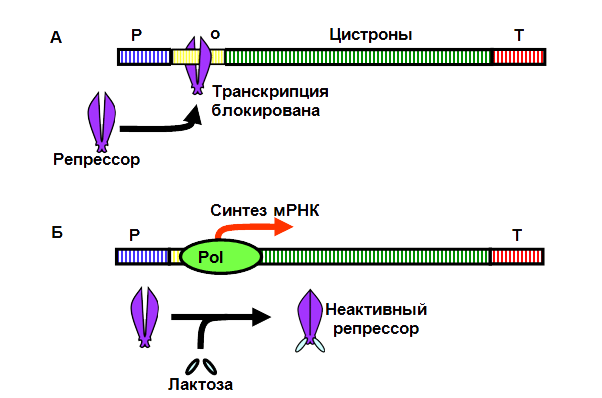
Рис. 12. Регуляция экспрессии лактозного оперона:
Pol – фермент РНК-полимераза; Р – промотор; о – оператор; T –терминатор
Кроме того, лактозный оперон может подвергаться позитивной индукции. Дело в том, что промотор лактозного оперона весьма слабый и даже после удаления белка-репрессора инициация транскрипции с него, без воздействия специального активирующего комплекса, почти не идёт. Активация не происходит до тех пор, пока в среде присутствует глюкоза. И это в полной мере отвечает потребностям самой клетки, поскольку глюкоза является более предпочтительным питательным субстратом и источником углерода. Поэтому нет никакой необходимости усиливать катаболизм «трудноперевариваемой» лактозы. Механизм позитивной индукции лактозного оперона требует, прежде всего, образования циклической формы аденозин-монофосфата (цАМФ). Реакцию его образования из АТФ катализирует фермент аденилат-циклаза. После этот с цАМФ взаимодействует белок CAP, в результате чего образуется активирующий комплекс цАМФ-CAP. Этот комплекс взаимодействует с ДНК в начале лактозного оперона и изменяет конформацию его промотора, что приводит к увеличению сродства промотора к РНК-полимеразе и усилению транскрипции оперона в десятки раз. Начинается усиленное потребление лактозы бактериальной клеткой.
Особенность данного механизма регуляции в том, что непосредственно глюкоза ингибирует аденилат-циклазу и активирует фермент фосфодиэстеразу (рис. 13). Последний катализирует перевод цАМФ в АМФ. Таким образом, при появлении глюкозы происходит разрушение активирующего комплекса, вызывающее значительное снижение уровня экспрессии лактозного оперона. Это пример позитивной репрессии.

Рис. 13. Регуляторное воздействие глюкозы на ферментативные процессы
 Франсуа Жакоб (фр. François Jacob)
Франсуа Жакоб (фр. François Jacob)
(1920 г.р.)
Французский микробиолог и генетик.
Родился в Нанси, Франция. После окончания лицея Карно в Париже, поступил в Сорбонну на медицинский факультет, выбрав в качестве профилирующего предмета хирургию. Учёба была прервана войной. В июне 1940 года вступил в «Свободные Французские вооруженные силы», базировавшиеся в Лондоне. Его приписали в качестве военного врача во 2-ю бронетанковую дивизию, направлявшуюся в Африку. В августе 1944 года он был тяжело ранен и семь месяцев провёл в госпитале. Награжден крестом Освобождения – одной из высших военных наград Франции. После войны Жакоб завершил образование и в 1947 году получил диплом врача. Полученные ранения не позволили ему стать хирургом и он в 1950 году начал работать в институте Пастера под руководством Андре Львова. Его первые научные исследования, проведённые в институте Пастера, посвящены генетическим механизмам, существующим в бактериях и бактериофагах (вирусах бактерий), и биохимическим эффектам мутаций. В 1965 году, совместно с А. Львовым и Ж. Моно, становится лауреатом Нобелевской премии по физиологии и медицине «за вклад в понимание фундаментальных процессов в живой материи, которые служат базисом таких явлений как адаптация, воспроизведение и эволюция»
Источник информации и фото: http://uzrf.ru
 Жак Люсье́н Моно (фр. Jacques Lucien Monod)
Жак Люсье́н Моно (фр. Jacques Lucien Monod)
(1910 – 1976)
Французский биохимик и микробиолог. Родился 9 февраля 1910 года в Париже в семье французского художника и американки из Милуоки, США. В 1917 году вместе с семьей переехал в Южную Францию, где провел бо́льшую часть своего детства. Своим интересом к биологии Жак был во многом обязан своему отцу, который был весьма эрудированным человеком и много читал Дарвина. Учился в Каннском лицее, в 1928 году поступил на факультет естественных наук в Париже, тогда еще не осознавая, что знания, даваемые там, отставали по крайней мере на двадцать лет от современной биологической науки.
В 1941 году получил докторскую степень в естественных науках. После периода преподавания на факультете наук в 1934 году и в Калифорнийском технологическом институте в 1936, Моно переходит в Пастеровский институт, где в то время освободилась должность директора лаборатории на факультете Андре Львова. В 1954 году Моно становится деканом факультета клеточной биохимии, а в 1959 профессором химии метаболизма в Сорбонне. В 1967 году получает должность профессора в Коллеж де Франс. В 1971 году Моно становится директором Пастеровского института.
Основные труды Моно посвящены изучению роста бактерий, индукции и репрессии ферментов, исследованию механизма регуляции синтеза белка у бактерий. Он разработал метод непрерывного культивирования бактерий. В 1958 предложил (совместно с Ф. Жакобом и А.М. Львовым) схему синтеза белка в бактериальной клетке. В 1961 высказал гипотезу о переносе генетической информации с ДНК на рибосомы при участии особой РНК – информационной (матричной; мРНК) и о переводе информации с языка нуклеотидов на язык аминокислот (синтез белка). Вместе с Ф. Жакобом сформулировал концепцию оперона - генного комплекса, детерминирующего синтез определенных белков (например, ферментов, участвующих в биосинтезе какого-либо метаболита). В своей широко известной биологической и философской работе Случайность и необходимость (Le Hasard et la Ncessit, 1970) Моно, основываясь на последних открытиях в области биохимии, утверждал, что все формы жизни – это результат случайных мутаций (случайность) и дарвиновского отбора (необходимость).
Источник информации: http://ru.wikipedia.org/ и http://dic.academic.ru
Источник фото: http://www.calend.ru/
Молекулярное строение лактозы