Транскрипция (рис. 40) (от лат. транскрипцио – переписывание) – биосинтез РНК на матрице (соответствующих участках) ДНК, осуществляющийся в клетках организма, – первый этап реализации генетической информации, в ходе которого последовательность нуклеотидов ДНК "переписывается" в нуклеотидную последовательность РНК. В процессе транскрипции на молекуле одной из полинуклеотидных нитей ДНК, служащей как бы матрицей (ДНК-матрица), при участии РНК-полимеразы синтезируются молекулы иРНК. При этом последовательность рибонуклеотидов иРНК в соответствии с комплементарностью азотистых оснований отражает последовательность дезоксирибонуклеотидов ДНК-матрицы. Происходит "считывание" генетического кода. При полном формировании молекула иРНК переходит из ядра в цитоплазму и прикрепляется к одной из рибосом, становясь матрицей для биосинтеза специфических белков – процесса трансляции. Транскрипция обратная – перенос ("переписывание") генетической информации с РНК на ДНК.
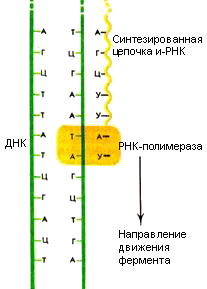
Рис. 40. Транскрипция
Первым этапом синтеза белка является транскрипция – процесс биосинтеза молекул РНК на участках ДНК – генах, который осуществляется специфическими ферментами (РНК-полимеразами). Матрицей для синтеза молекул РНК служит кодогенная цепь ДНК. Одни гены содержат информацию о последовательностях аминокислот в полипептидных цепочках и, следовательно, иРНК, другие – о последовательностях нуклеотидов в молекулах рРНК или тРНК.
При синтезе иРНК происходит переписывание генетической информации с гена, в котором записана информация о последовательности аминокислот в одной полипептидной цепочке, на иРНК. Считывание всегда идет в одном направлении от 3'-концу к 5'-концу матрицы.
Большинство эукариотических генов содержат в своей нуклеотидной последовательности протяженные нетранслируемые вставки, которые прерывают соответствие между нуклеотидной последовательностью остальных участков гена и аминокислотной последовательностью полипептида, кодируемого этим геном. Такие нетранслируемые участки ДНК в генах называют интронами, тогда как участки гена, кодирующие аминокислотную последовательность полипептида, называют экзонами. Интронные последовательности РНК специфически выщепляются из каждого РНК-транскрипта в ходе его превращения в "зрелую" молекулу мРНК, способную участвовать в синтезе белка. Поскольку экзоны после вырезания интронов соединяются друг с другом в стык и образуют единое целое, процессинг РНК, протекающий по такой схеме, получил название сплайсинг (от англ. splice – сращивать). В тех случаях, когда ген содержит несколько интронов, важно, чтобы экзоны соединялись в правильной последовательности, а не случайным образом (рис. 41).
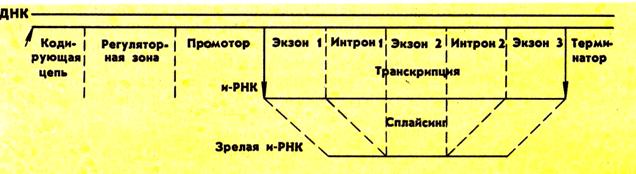
Рис. 41. Структура гена
В некоторых случаях в клетке осуществляются несколько детерменированных альтернативных сплайсингов, что приводит к образованию из одного и того же РНК-транскрипта нескольких отличающихся друг от друга молекул иРНК, кодирующих различные белки. Примером может служить формирование многообразия антител в лимфоцитах. Возможно также, что альтернативный сплайсинг происходит в процессе клеточной дифференцировки. Таким образом, само существование сплайсинга придает клетке дополнительную генетическую гибкость. Известны случаи, когда интрон одного гена может быть экзоном другого гена, или когда интрон самостоятельно кодирует небольшой регуляторный белок. Одно из предположений связывает происхождение интронов у эукариот с появлением возможности комбинирования экзонов ("мини-генов"), каждый из которых кодирует функционально значимый белковый домен, что приводит к возникновению в ходе эволюции новых генов, кодирующих белки с новыми полезными свойствами.
Только около 5% общей массы РНК-транскриптов покидает клеточное ядро.