Расшифрована впервые в 1953 г. Уотсоном и Крик (Watson, Crick).
Вторичная структура ДНК представляет собой свернутые в спираль две комплементарно взаимодействующие и антипараллельные полинуклеотидные цепи. Образование вторичной структуры НК возможно вследствие проявления эффектов комплементарности и стэкинг-взаимодействий. Очень часто наблюдаются двунитевые спирализованные молекулы ДНК, замкнутые в кольцо с ковалентно связанными концами. Они не имеют разрывов у каждой в отдельности полинуклеотидной цепи. Подобные кольцевые ДНК, как правило, суперспирализованы, то есть кольцо дополнительно закручено в спираль. Суперспирализация – правило, а не исключение, при условии отсутствия разрывов в фосфодиэфирных связях полинуклеотидной цепи.
Комплементарность – последовательность нуклеотидов в одной цепи автоматически определяет строго соответствующую ей последовательность нуклеотидов в комплементарной ей цепи. Так, азотистое основание аденин (А) всегда взаимодействует только с комплементарным ему азотистым основанием тимин (Т) в молекулах ДНК.
Одновременно азотистые основания гуанин (Г) одной цепи взаимодействует только с комплементарными им азотистыми основаниями цитозин (Ц) другой цепи (как в ДНК, так и в РНК). Комплементарность оснований обеспечивается системой водородных связей. В молекулах РНК, имеющих, в основном, однонитевую структуру, на отдельных участках азотистые основания А взаимодействуют с комплементарными им азотистыми основаниями урацил (У).
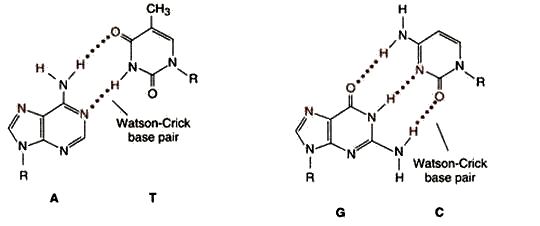
Аналогично происходит взаимодействие в процессах транскрипции, когда на матрице ДНК синтезируется молекула РНК (матричная, транспортная и т.д.), и наоборот, когда при участии реверс-транскриптазы происходит синтез кДНК на матрице РНК.
Стэкинг-взаимодействия – особого рода (Ван-дер-Ваальсовы) взаимодействия между выложенными в стопку (как монеты) друг над другом азотистыми основаниями.