Основные функциональные комплексы ЭТЦ
Изучение тонкой структуры мембран тилакоидов с помощью методов электронной микроскопии показало, что компоненты ЭТЦ фотосинтеза сгруппированы в комплексы. Структуру комплексов определяют входящие в их состав молекулы белков, а функциональные характеристики комплексов – пигменты, способные к окислительно-восстановительным превращениям, кофакторы, катионы и анионы. Латеральное распределение комплексов в мембранах имеет свои особенности, в зависимости от структурной организации внутренних мембран хлоропластов и их функционального состояния, например в условиях освещения и темноты. Отношение «белок/липид» на контактирующих участках тилакоидов гран выше, чем на участках тилакоидов стромы. Изменение положения комплексов в плоскости мембран тилакоидов происходит при условии, если липиды находятся в жидкокристаллическом состоянии. В механизме латерального перемещения светособирающего хлорофилл-а/в-белкового комплекса участвуют ионы магния, фосфат-анион и фермент киназа.
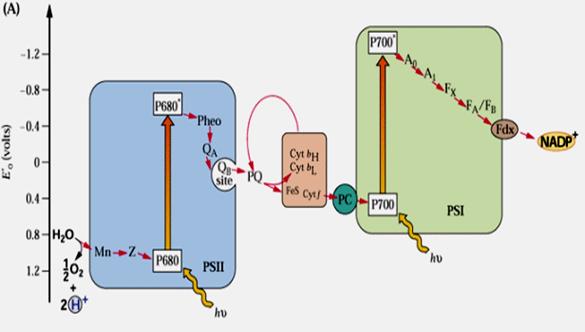
В цепи транспорта электрона выделяют пять белковых комплексов, дифференцированных структурно и функционально: I – комплекс ФС2, включающий в свой состав белки пигментной антенны, тесно связанные с РЦ, и комплекс фотоокисления воды; II – комплекс ФС1, также включающий в свой состав белки пигментной антенны и функционально связанный с ферредоксином и ферредоксин-НАДФ-редуктазой; III – светособирающий хлорофилл-а/б-белковый комплекс, ответственный за образование гран и регуляцию передачи энергии возбуждения между двумя фотосистемами; IV – цитохром b6/f комплекс с Fe-S-центром Риске; V – CF0-CF1-комплекс, представляющий собой АТФазную систему. Комплекс V локализован в тех частях мембраны, которые контактируют со стромой. Комплекс III расположен в зоне «слипания» гран. Комплекс IV участвует в Q-цикле.
Системы фотоокисления воды и выделения кислорода при фотосинтезе
Комплекс фотоокисления воды интегрирован в белок в составе ФС2. Реакции фотоокисления воды протекают на внутренней стороне мембран тилакоида. Реакцию фотоокисления воды можно записать как:
2Н2О →(4 кванта света)→ 4 Н+ + 4 ē + О2.
Для осуществления этой реакции необходимо согласовать образование окислительных эквивалентов в реакционном центре, происходящее как одно электронный процесс при поглощении каждого кванта света, с четырехэлектронным окислением воды до молекулярного кислорода. Кроме того, необходимо предотвратить взаимодействие промежуточных продуктов фотоокисления воды с другими молекулами и обеспечить равномерное распределение свободной энергии по последовательным стадиям фотоокисления. В настоящее время установлены следующие черты процесса фотоокисления воды. Четыре электрона отщепляются от двух молекул воды последовательно в результате четырех актов поглощения энергии света в одном реакционном центре (альтернатива кооперативной гипотезе). Молекулярный кислород образуется после удаления четырех электронов из воды, но ионы водорода высвобождаются друг за другом. Эти факты легли в основу циклической модели А. Жолио и Б. Кока (1970, 1975).
Переход из любого S-состояний в следующее происходит в момент, когда фотоокисленный компонент «Z» в составе РЦ ФС2 получает электрон от комплекса фотоокисления воды. После выключения света комплексы в состояниях S3 и S2 в ходе обратных реакций переходят в устойчивое состояние S1. Существование S-цикла подтверждают эксперименты, в которых послтемнового адаптационного периода в ходе импульсного освещения изолированных хлоропластов наблюдают повышение выхода кислорода при третьей вспышке в первом цикле, а затем при каждой четвертой вспышке в последующих циклах. Выделение 4-х протонов в S-цикле не является строго синхронизированным с окислительно-восстановительными превращениями компонента «Z». По одному протону выделяется на этапах S0 → S1 и S2 → S3, по два протона – на этапе S4 → S4.
Состав каталитического центра определен как Mn4O4Ca. В качестве кофактора реакции фотоокисления воды выступают ионы хлора. Центр фотоокисления воды блокируют тепловая обработка при температуре выше 40 °С, трис-буфер в концентрации 0,4 М, гидроксиламин.
Типы функциональной организации ЭТЦ: нециклический, циклический и псевдоциклический потоки электронов и фотофосфорилирование
Путь электрона от воды к НАДФ+ с участием двух фотосистем получил название нециклического транспорта электронов. В результате образуются молекулярный кислород, восстановленная форма НАДФН+Н и мембранный электрохимический градиент ионов водорода (между стромой и пространством внутри тилакоида). Кроме нециклического потока электронов при определенных условиях может происходить циклический транспорт электронов (с участием одной ФС1 или одной ФС2) и псевдоциклический (с участием обеих фотосистем).
Циклический транспорт с участием ФС2 защищает РЦ ФС2 от фоторазрушения при высокой интенсивности света, циклический транспорт с участием ФС1 способствует созданию градиента ионов водорода и тем самым участвует в поддержании необходимого энергетического состояния мембраны.
Псевдоциклический транспорт необходим в первый момент освещения после темнового периода для активации ионного транспорта и перехода фотосинтетического аппарата в функциональное состояние.
В эксперименте с изолированными хлоропластами можно моделировать работу нециклического транспорта электронов на участке от воды до НАДФ+(ФС2+ФС1), нециклического транспорта от воды к искусственному акцептору электронов, функционирующему между двумя фотосистемами (ФС2), циклического с участием ФС1 и псевдоциклического электронного транспорта (ФС2+ФС1).